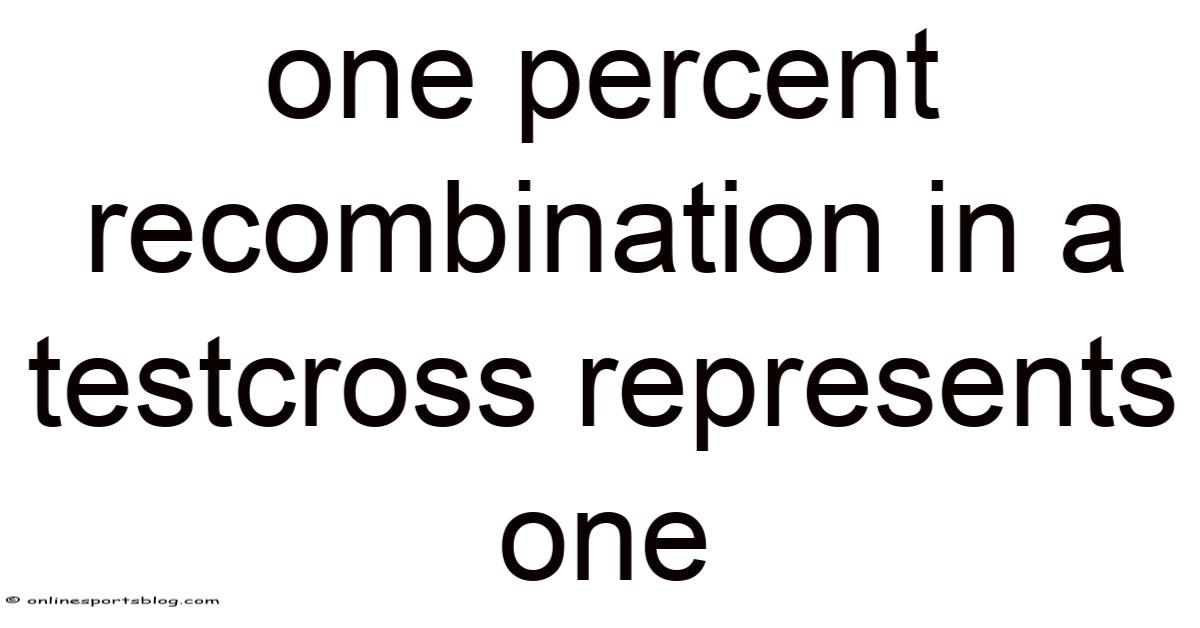
Introduction
One percent recombinationin a testcross represents one centimorgan, a measure of genetic distance that indicates the likelihood of a crossover event between two genes. So this seemingly small value carries profound implications for mapping genomes, understanding inheritance patterns, and predicting the outcomes of breeding programs. In classical genetics, a testcross—where an individual of unknown genotype is mated with a homozygous recessive partner—provides a clear window into the segregation of alleles and the frequency of recombination. When the observed recombination frequency is exactly one percent, it tells us that the two loci under study are separated by a distance of one map unit, or one centimorgan (cM). The following article walks you through the conceptual foundation, the practical steps involved in obtaining this value, the underlying biological mechanisms, and answers to frequently asked questions, all while maintaining a clear, engaging, and SEO‑friendly narrative Surprisingly effective..
Understanding the Testcross
A testcross is a genetic cross between an individual with a dominant phenotype (but an ambiguous genotype) and a homozygous recessive individual. The offspring phenotypes reveal the hidden genotype of the dominant parent. If the dominant parent is heterozygous at a particular locus, the expected ratio of dominant to recessive phenotypes in the progeny is 1:1. When two loci are considered, the proportion of recombinant phenotypes (those that display a new combination of traits) compared to parental phenotypes reflects the recombination frequency between them Easy to understand, harder to ignore..
Quick note before moving on.
Steps to Perform a Testcross
- Select Parental Strains
- Choose a true‑breeding recessive line (e.g., aa) and a dominant line whose genotype at the target loci is unknown (e.g., A_).
- Set Up the Cross
- Mate a heterozygous individual (AaBb) with the recessive partner (aabb). This produces a testcross progeny where each allele segregates independently.
- Count Offspring Phenotypes
- Observe a large sample (ideally >1000 individuals) to minimize sampling error. Record the numbers of parental (AB/ab) and recombinant (Ab/aB) phenotypes.
- Calculate Recombination Frequency
- Use the formula:
[ \text{Recombination Frequency (%)} = \frac{\text{Number of Recombinants}}{\text{Total Offspring}} \times 100 ] - If the result is 1%, the loci are one centimorgan apart, meaning that one crossover event occurs per 100 meioses on average.
- Use the formula:
Why Sample Size Matters
Recombination events are rare, especially at small distances. A larger sample size reduces stochastic variance, giving a more accurate estimate of the true recombination frequency. In practice, geneticists aim for at least 10 × the expected number of recombinants to achieve reliable statistics Took long enough..
Scientific Explanation
What Is Recombination?
Recombination, or crossing over, occurs during prophase I of meiosis when homologous chromosomes exchange segments. This shuffling creates new allele combinations, increasing genetic diversity. The physical distance between two genes on a chromosome correlates with the probability that a crossover will separate them.
One Percent = One Centimorgan
In genetic mapping, a centimorgan (cM) is defined as the distance that yields a 1 % recombination frequency. Because of this, a one percent recombination in a testcross represents one centimorgan. This unit is not a physical length (like a base pair) but a functional measure of how often a crossover separates the markers. In most organisms, 1 cM approximates 1 million base pairs, though this ratio varies across species and chromosomal regions.
Some disagree here. Fair enough It's one of those things that adds up..
Molecular Basis
At the molecular level, recombination is mediated by the formation of double‑strand breaks and the subsequent repair via homologous recombination pathways. The spliceosome, recA‑like proteins, and merosome structures coordinate the exchange. The frequency of these events is influenced by:
- Chromosome architecture – euchromatin vs. heterochromatin.
- Hotspot activity – specific DNA sequences that boost crossover rates.
- Sex-specific differences – in many species, recombination is higher in females.
Implications for Gene Mapping
When researchers construct a genetic map, they order markers based on recombination frequencies. A one percent recombination places two markers only one step apart, implying tight linkage and limited shuffling of alleles across generations. This tight linkage can be advantageous for:
- **Marker‑ass
The study of parental and recombinant phenotypes provides critical insight into the genetic architecture of organisms under investigation. Think about it: by examining the counts of AB/ab and Ab/aB types, scientists can discern how effectively chromosomes shuffle during meiosis. This understanding not only refines the mapping of genes but also informs strategies for breeding or genetic engineering.
Quick note before moving on.
Calculating the recombination frequency from observed data strengthens the link between molecular mechanisms and phenotypic variation. Since recombination rates directly influence how alleles are distributed, accurately determining these percentages allows researchers to predict inheritance patterns and evolutionary outcomes.
Understanding these processes also highlights the importance of sample size; larger datasets enhance reliability, especially when dealing with subtle genetic changes. When all is said and done, this analysis reinforces the dynamic nature of the genome and the constant interplay between genetic recombination and selection pressures That's the whole idea..
To wrap this up, mapping parental and recombinant forms is more than a statistical exercise—it is a foundational step in unraveling the complexity of heredity and adaptation. Recognizing the significance of even small recombination percentages deepens our appreciation of genetic diversity Easy to understand, harder to ignore..
Conclusion: naturally integrating phenotype data with recombination calculations offers a clearer picture of genetic linkage and diversity, underscoring the necessity of precise measurement for meaningful scientific conclusions Turns out it matters..
This integration of molecular mechanisms and phenotypic outcomes underscores the interconnectedness of genetic processes. Recombination, while a molecular event, manifests as observable patterns in offspring, bridging the gap between cellular biology and evolutionary biology. Take this case: the precise mapping of recombination hotspots through phenotypic analysis informs efforts to manipulate crop genomes for desired traits, such as drought resistance or pest resistance in agriculture. Similarly, in human genetics, understanding recombination frequencies aids in identifying chromosomal abnormalities and refining linkage studies for disease-associated genes.
The dynamic nature of recombination also has profound implications for evolutionary theory. And regions of the genome with high recombination rates, such as those near hotspots, exhibit greater genetic diversity, accelerating adaptation to environmental changes. Conversely, tightly linked regions, constrained by low recombination, may preserve co-adapted gene complexes critical for survival. This duality highlights recombination as both a driver of innovation and a conservator of functional genomic architecture Less friction, more output..
So, to summarize, the study of parental and recombinant phenotypes is a cornerstone of genetic research, offering insights into the molecular, cellular, and evolutionary dimensions of heredity. In practice, by quantifying recombination frequencies and integrating them with phenotypic data, scientists can unravel the complexities of genetic linkage, refine mapping techniques, and harness these principles for applications ranging from personalized medicine to sustainable agriculture. The bottom line: recombination remains a testament to the genome’s adaptability—a mechanism that ensures both stability and diversity across generations.
Continuing without friction from the established themes of recombination's role in evolution and its practical applications, the field is rapidly advancing through technological innovations. High-throughput sequencing now allows for the precise identification of recombination breakpoints across entire genomes in large populations, revealing patterns previously obscured by limited sampling. This granular data enables the construction of ultra-dense genetic maps, pinpointing recombination hotspots with unprecedented accuracy and uncovering the influence of chromatin structure, DNA sequence motifs, and even epigenetic marks on recombination rates. What's more, the integration of CRISPR-based gene editing technologies allows researchers to deliberately introduce or suppress recombination events at specific genomic loci, providing direct experimental validation of hypotheses about linkage and trait inheritance derived from mapping studies.
The interplay between recombination and selection also manifests in more nuanced ways. While recombination generally breaks down linkage disequilibrium (LD), the very regions under strong selective pressure, such as those involved in host-pathogen co-evolution or adaptation to extreme environments, can exhibit unique recombination dynamics. These regions might show suppressed recombination to maintain co-adapted alleles or, conversely, elevated recombination to generate novel combinations beneficial under fluctuating selective pressures. Understanding this context-dependent response is crucial for interpreting patterns of genetic diversity and for predicting how populations might respond to future environmental challenges or disease pressures. Because of this, modern recombination mapping increasingly incorporates population genomic data alongside phenotypic information to disentangle the effects of selection, demography, and recombination itself.
Honestly, this part trips people up more than it should.
To wrap this up, the meticulous mapping of parental and recombinant forms, integrated with advanced molecular techniques and phenotypic analysis, has evolved into a powerful cornerstone of modern genetics. It provides a fundamental lens through which we decipher the nuanced dance of DNA exchange, revealing how recombination shapes genetic diversity, fuels adaptation, and constrains evolutionary pathways. It underscores that the genome is not a static blueprint but a dynamic, responsive entity, where the shuffling of genetic information is a continuous, essential process ensuring resilience and innovation across generations. From enabling precision breeding in agriculture to refining the search for disease genes and illuminating the mechanisms of speciation, the study of recombination bridges the molecular realm of DNA mechanics with the grand narrative of life's evolution. Future research, leveraging ever more sophisticated tools, promises to further unravel the complex regulation and profound consequences of this fundamental biological mechanism.